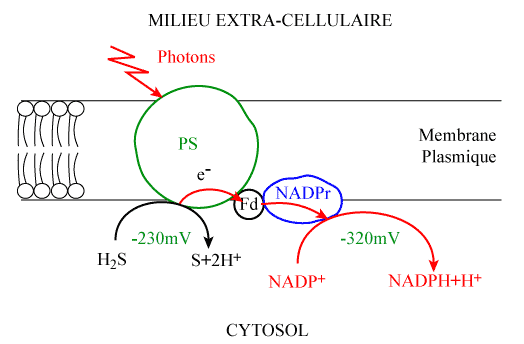
Lors de l'apparition des premières formes de vie, l'atmosphère était exempte d'oxygène et donc de nature essentiellement réductrice. Les premières cellules procaryotes étaient donc anaérobies et donc capables de produire leur énergie sans faire appel à l'oxygène. Les premières voies de conversion de l'énergie utilisées par les cellules sont vraisemblablement celles de la fermentation qui permettaient d'utiliser les (rares) molécules organiques en absence d'oxygène. La raréfaction des molécules organiques a favorisé la sélection de bactéries photoautotrophes, possédant des chaînes de transport d'électrons comportant un photosystème (PS I) et capable de synthétiser des molécules organiques. Ainsi, les sulfobactéries réalisent un type de photosynthèse dans laquelle le donneur d'e- est H2S (figure 7). Une seconde étape de l'évolution des chaînes photosynthétiques, réalisées par les cyanobactéries, voit apparaître un deuxième photosystème (PS II). Le niveau d’énergie ainsi atteint permet d’utiliser comme donneur d’électrons une molécule dont le potentiel de réduction est plus élevé que H2S : l’eau (figure 5).
| Figure 7. Photosynthèse chez une sulfobactérie |
|
Les cyanobactéries ont donc produit de l'oxygène qui s'est accumulé
dans l'atmosphère qui, de réducteur, est devenu oxydant. Ce changement
favorise non seulement les organismes capables de tolérer la présence
d'oxygène dans l'atmosphère mais surtout ceux qui sont capables
de l'utiliser pour oxyder les molécules organiques qui, par ailleurs,
s'accumulent par suite de l'activité photosynthétique. Ces organismes
sélectionnés par l'évolution sont les bactéries
hétérotrophes possédant des chaînes respiratoires
puisque la respiration est beaucoup plus efficace que la fermentation.
| Figure 8. Transport cyclique d'électrons chez une bactérie pourpre |
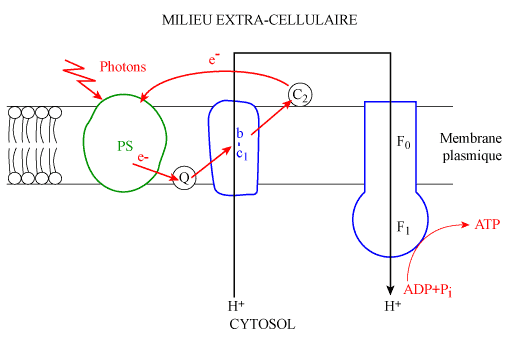 |
Cette adaptation des procaryotes aux changements de composition de l'atmosphère
traduit donc l'évolution des chaînes de transport des électrons.
La comparaison de ces dernières
( voir figures 3 et 5 page 10.3 et figures 7 et 8 ci-dessus) fait apparaître
des analogies frappantes. Ainsi, la portion de la chaîne, présente
chez les mitochondries et les chloroplastes actuels, et constituée d'une
quinone cédant des électrons à un complexe de cytochromes
pompant des électrons à travers la membrane, existait déjà
chez des bactéries anaérobies (figure 8), il y a plus de 3 milliards
d'années.
Ainsi, par exemple, l'ADN mitochondrial des mammifères est une molécule circulaire de 5,5 µm (17 000 pb). Il comprend 37 gènes : gènes des 2 ARN ribosomiques et de 22 ARNt et seulement 13 gènes codant des polypeptides appartenant aux complexes de la chaîne respiratoire. La majorité des protéines mitochondriales est donc codée par le génome nucléaire et synthétisée dans le cytosol, ce qui justifie le terme de semi-autonomie génétique.
Au contraire, toute mitochondrie, ou tout plaste, résulte de la division binaire d'un organite préexistant, comme l'a montré Lucke (1963) pour les mitochondries de Neurospora crassa. Cette division est précédée d'une réplication de l'ADN mitochondrial ou chloroplastique. Cette synthèse est asynchrone de celle de l'ADN nucléaire, et donc totalement indépendante de cette dernière.
Ce processus de biogenèse par bipartition est tout à fait en accord avec le fait que ces organites sont doués de continuité génétique et d'une certaine autonomie sur le plan de l'information génétique.
De nombreux points communs entre mitochondries, chloroplastes et bactéries peuvent être relevés : nature des lipides membranaires, parenté entre ribosomes bactériens et mitoribosomes et surtout plastoribosomes, sensibilité aux antibiotiques, structure des gènes, similitude des chaînes de transport d'électrons (voir paragraphe 5.4.1). L'existence à l'heure actuelle de bactéries symbiotiques vivant dans le cytoplasme de cellules eucaryotes vient également à l'appui de la théorie endosymbiotique.
Selon la théorie de l'endosymbiose en série, le scénario serait le suivant. Une première endosymbiose d'une bactérie hétérotrophe aérobie, proche des bactéries pourpres actuelles est à l'origine des mitochondries présentes dans toutes les cellules eucaryotes. Elle a été suivie d'une seconde endosymbiose impliquant une cyanobactérie qui est à l'origine des chloroplastes des cellules eucaryotes photosynthétiques.
Les concepts actuels de la biologie moléculaire permettent d'imaginer les modalités du processus de transformation du symbionte en organite semi-autonome. En effet, les développements du génie génétique montrent que de l'ADN exogène peut être incorporé aux chromosomes d'une cellule eucaryote (transgenèse). Un lent processus de transfert de gènes s'est opéré, depuis le génome des bactéries symbiotiques vers les chromosomes nucléaires des cellules hôtes. Ces dernières synthétisent maintenant la plus grande partie des molécules constitutives des mitochondries et des plastes, auxquels ces molécules doivent parvenir par des systèmes d'adressage spécifiques.