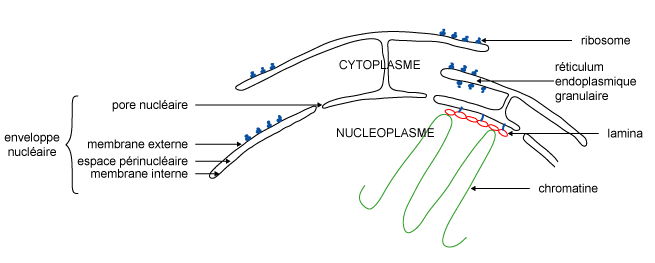
Figure 1
L'enveloppe nucléaire est constituée de 2 membranes séparées par un intervalle de 20 à 40 nm, l'espace périnucléaire. Elle est percée de pores nucléaires. La membrane externe est en continuité avec la membrane du réticulum endoplasmique. L'enveloppe nucléaire est en fait une différenciation locale de ce dernier (figure 1).
|
Figure 1 |
La membrane interne est doublée d'une couche protéique, la lamina,
qui s'interrompt au niveau des pores. La lamina est constituée de l'agencement
de protéines, les lamines qui forment un réseau sur lequel est
ancré la chromatine.
Les pores nucléaires ne sont pas de simples perforations dans l'enveloppe.
Un complexe protéique d'une masse moléculaire de 1,2 x 108
Da comprenant jusqu'à 200 peptides est associé à chaque
pore (figure 2).
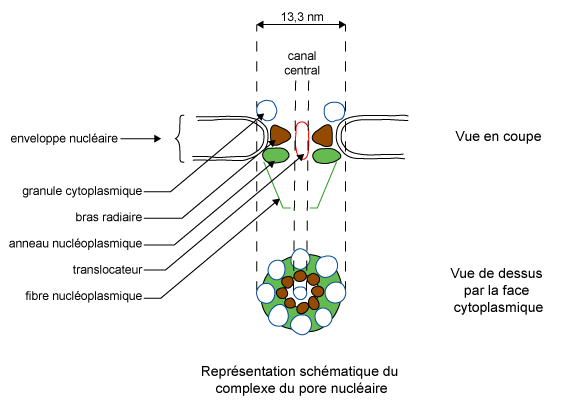 |
Figure 2 : Représentation
schématique du complexe du pore nucléaire |
Ce matériel laisse ouvert un canal central. Ce dernier est souvent obstrué par un "granule central" parfois appelé translocateur. Pendant longtemps, les observations en microscopie électronique de passage de matériel à travers les pores nucléaires sont restées difficilement interprétables. Par contre, les études de perméabilité de l'enveloppe nucléaire ont montré que les pores sont impliqués dans les échanges entre noyau et cytoplasme. Les résultats indiquent que les molécules indéformables d'une masse moléculaire inférieure à 60-70 kDa diffusent plus ou moins rapidement par les pores nucléaires et sont compatibles avec l'existence de canaux aqueux de 9 nm de diamètre. Les macromolécules ou les édifices macromoléculaires de masse moléculaire supérieure doivent subir une déformation. Ceci a été clairement observé dans le cas des cellules de glande salivaire de Chironome (figure 3).
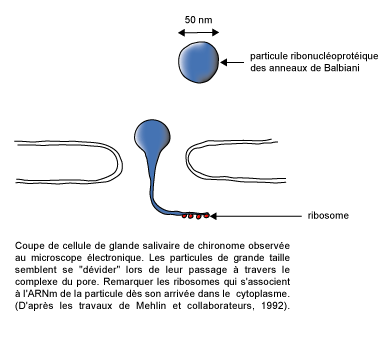 |
| Figure 3 : Coupe de cellule de glande salivaire observée au microscope électronique. Les particules de grande taille semblent se "dévider" lors de leur passage à travers le complexe du pore. Remarquer les ribosomes qui s'associent à l'ARNm de la particule dès son arrivée dans le cytoplasme. (D'après les travaux de Mehlin et collaborateurs, 1992)
|
Par ailleurs, le transport de beaucoup de molécules par l'intermédiaire
des pores nucléaires ne résulte pas d'une simple diffusion mais
fait intervenir -comme pour les autres compartiments cellulaires- un mécanisme
"d'adressage" dans lequel certains polypeptides du complexe du pore
jouent un grand rôle.