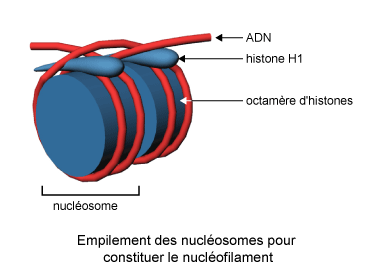
Figure 4 : Empilement des nucléosomes pour constituer le nucléofilament
Les techniques de fractionnement cellulaire permettent d'obtenir une fraction pure de noyaux à partir d'un homogénat cellulaire. L'action des ultrasons rompt l'enveloppe nucléaire. Trois sous-fractions peuvent alors être isolées :
Le dosage chimique de la sous-fraction chromatine montre qu'elle renferme 4 constituants principaux :
L'ADN et les histones forment un complexe (nucléohistones) difficile à dissocier. Les histones sont des protéines basiques, donc riches en lysine et en arginine. Il existe 5 types de molécules d'histones qui se différencient notamment par leurs teneurs respectives en lysine et en arginine : H1, H2a, H2b, H3 et H4.
L'extraction de la chromatine par une solution de faible force ionique donne,
en microscopie électronique une image appelée "collier de
perles". Elle montre que la chromatine est constituée d'unités
répétitives de 11 nm de diamètre, appelées nucléosomes
(les "perles" du collier), reliées entre elles par un filament
de 2,5 nm de diamètre : le lien internucléosomique, qui correspond
en fait à la molécule d'ADN
(figures 4 et 5).
Chaque nucléosome est constitué d'un cœur de 8 molécules
d'histones autour duquel est enroulée la molécule d'ADN sur une
longueur de 146 paires de bases (figure 4).
|
|
Figure 4 : Empilement des nucléosomes pour constituer le nucléofilament |
Très utile pour mettre en évidence la structure nucléosomique
de la chromatine, l'extraction en solution de faible force ionique ne donne
néanmoins pas une idée correcte de la structure de la chromatine
native. En réalité, les liens internucléosomiques ne sont
pas étirés et les histones H1 permettent la juxtaposition des
nucléosomes pour former une fibre cylindrique d'un diamètre constant
de 10 nm : le nucléofilament (figure 4).
L'extraction à concentration saline physiologique permet de constater
que la chromatine native se présente sous l'aspect d'une fibre de 30
nm de diamètre, la fibre chromosomique, résultant de l'enroulement
du nucléofilament, vraisemblablement sous forme d'un solénoïde
(figure 5).
 |
Figure 5 : Organisation de la chromatine |
Cette architecture moléculaire de la chromatine constitue un premier niveau de compaction de la molécule d'ADN. En effet, un noyau interphasique de quelques micromètres de diamètre doit pouvoir renfermer, par exemple dans l'espèce humaine, 46 chromosomes contenant chacun une molécule d'ADN d'une longueur moyenne de 5 cm. La configuration nucléosomique de la chromatine réduit la longueur de la structure selon un facteur 7, l'enroulement du nucléofilament en fibre chromosomique de nouveau selon un facteur 6, ce qui réduit l'encombrement de la molécule d'ADN de 5 cm à environ 1,2 mm.
Dans le noyau interphasique, les fibres chromosiques forment un réseau
continu plus ou moins condensé dans lequel il est impossible d'individualiser
les différents chromosomes. Ces derniers ne sont identifiables en tant
que tels que pendant la division cellulaire. Le chromosome métaphasique
est la forme sous laquelle la compaction de la fibre chromosomique est la mieux
connue.
La métaphase de mitose est la période du cycle cellulaire pendant
laquelle la condensation de la chromatine est maximale. Le chromosome métaphasique
est constitué de 2 chromatides associées au niveau du centromère
(figure 6).
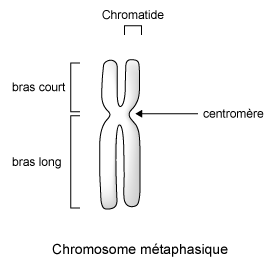 |
Figure 6 : Chromosome métaphasique |
La surface d'un tel chromosome isolé et observé au microscope électronique à balayage révèle des microconvules qui sont la forme ultime de condensation de la chromatine (figure 7).
 |
Figure 7 : Structure du chromosome |
Une déshydratation ménagée du chromosome provoque le déploiement
des microconvules en boucles formées d'une fibre de 30 nm de diamètre,
il s'agit de la fibre chromosomique.
Lorsque le chromosome subit un traitement qui élimine les histones (figure
7), le microscope électronique révèle que l'ADN, complètement
déployé par suite de la perte de la structure nucléosomique,
forme un halo autour d'un échafaudage (ou charpente) de protéines
non histones, ayant la forme spécifique du chromosome métaphasique.
L'observation des préparations à plus fort grossissement montre
que l'ADN forme des boucles de 10 à 30 µm de circonférence
dont les deux extrémités sont insérées côte
à côte dans la charpente protéique. Les observations réalisées
en microscopie confocale laissent penser que la charpente protéique de
chaque chromatide forme une hélice régulière.
Ainsi, l'enroulement de la fibre chromosomique sur l'échafaudage protéique
de la chromatide introduit une compaction supplémentaire. La fibre chromosomique
de 1,2 mm est empaquetée en un chromosome de 5 µm de longueur.
Dans un chromosome, L'ADN est donc au total compacté selon un facteur
de l'ordre de 10 000. Dans le noyau interphasique, la compaction est de l'ordre
de 680. La fibre chromosomique semble alors être insérée
sur la matrice nucléaire dont au moins certaines protéines seraient
identiques à celles de la charpente chromosomique.
Le degré de compaction de l'ADN n'est pas le même dans toute la
chromatine du noyau interphasique. On distingue d'une part la chromatine condensée,
assimilée à l'hétérochromatine, et d'autre part
la chromatine dispersée, assimilée à l'euchromatine, qui
serait active du point de vue de la transcription.