Figure 1
Formation d'un di(désoxy)ribonucléotide
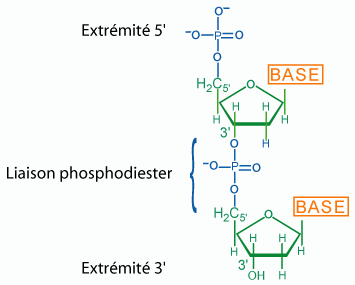
L’ADN est un polymère non ramifié de 4 désoxyribonucléosides monophosphates dont les bases sont respectivement : l’adénine, la guanine, la cytosine et la thymine (voir chapitre 2).
La polymérisation s’effectue par estérification de la fonction alcool 3’ du désoxyribose d’un nucléotide par l’acide phosphorique du nucléotide suivant. Les liaisons entre les nucléotides sont donc des liaisons phospho-diesters entre le carbone 3’ d’un nucléotide et le carbone 5’ du nucléotide suivant (figure 1).
Figure 1 |
|
Un brin d’ADN, comme d’ailleurs toute molécule d’acide nucléique, présente donc à une de ses extrémités une fonction acide phosphorique non engagée dans une liaison phospho-diester (extrémité 5’P), et de même à l’autre extrémité une fonction alcool libre (extrémité 3’OH). Ceci défini l’orientation des molécules d’acides nucléiques (Cf celle des protéines, voir chapitre 2) et on écrit, par convention internationale, la séquence des acides nucléiques en indiquant les initiales des bases en partant de l’extrémité 5’P à gauche, par exemple :
5’P-AGT ……………… GCT-3’OH
ou, plus simplement :
5’-AGT ..……………. GCT- 3’
Lorsque les extrémités 5’ et 3’ ne sont pas expressément indiquées :
AGT ……………… GCT
il est donc entendu, par convention, que l’extrémité 5’ est à gauche, l’extrémité 3’ à droite.
A partir de données physico-chimiques, et notamment les résultats de diffractions des rayons X (réalisées par Rosalind Franklin), Watson et Crick ont proposé en 1953 un modèle bicaténaire hélicoïdal : la « double hélice d’ADN » qui est constituée de deux chaînes polynucléotidiques reliées par des liaisons hydrogène entre les bases azotées (figure 2).
 |
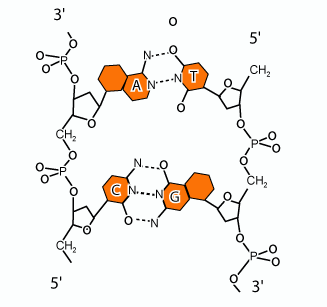 |
Figure 2 |
|
Les caractéristiques importantes de la structure tridimensionnelle de la molécule d’ADN sont les suivantes :
Note : on mesure la longueur des molécules d’ADN en paires de bases
(pb) pour bien marquer le caractère bicaténaire de la molécule
et le fait que ce sont les bases azotées des nucléotides qui jouent
le rôle essentiel dans l’appariement des deux brins.